The purpose of this post is to demonstrate some very beautiful (I think!) mathematics that arises form Darwinian evolutionary theory. It is a real shame that most courses and discussions dealing with evolution never introduce any type of mathematical formalism which is very strange, since at the most fundamental levels, evolution must also be governed by quantum mechanics and electromagnetism, from which chemistry and biochemistry arise via top-down and bottom-up causation. See this article by George Ellis for more on the role of top-down causation in the universe and the hierarchy of physical matter. Indeed, my personal belief is that if some biologists and evolutionary biologists like Dawkins, Coyne, and others took the time to explain evolution with some modicum of mathematical formalism to properly describe the underlying mechanics instead of using it as an opportunity to attack religious people, the world would be a much better place, and the dialogue between science and religion would be much more smooth and intelligible.
In this post today, I will describe some formalism behind the phenomena of prebiotic evolution. It turns out that there has been a very good book by Claudius Gros and understanding evolution as a complex dynamical system (dynamical systems theory is my main area of research), and the interested reader should check out his book for more details on what follows below.
We can for simplicity consider a quasispecies as a system of macromolecules that have the ability to carry information, and consider the dynamics of the concentrations of the constituent molecules as the following dynamical system:

where 
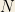
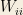

From this, we can consider the following catalytic reaction equations:

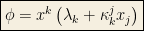
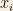



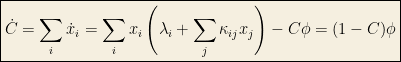
Clearly:

that is, this quick calculation shows that the total concentration 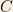
Let us consider now the case of homogeneous interactions such that

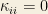

which leads to

which becomes

This is a one-dimensional ODE with the following invariant submanifolds:
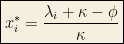
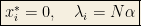
With homogeneous interactions, the concentrations with the largest growth rates will dominate, so there exists a 
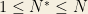
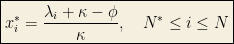

The quantities
![\boxed{1 = \frac{\alpha}{2\kappa} \left[N(N+1) - N^*(N^* - 1)\right] + \left[\frac{\kappa - \phi}{\kappa}\right] \left(N + 1 - N^*\right)}](https://s0.wp.com/latex.php?latex=%5Cboxed%7B1+%3D+%5Cfrac%7B%5Calpha%7D%7B2%5Ckappa%7D+%5Cleft%5BN%28N%2B1%29+-+N%5E%2A%28N%5E%2A+-+1%29%5Cright%5D+%2B+%5Cleft%5B%5Cfrac%7B%5Ckappa+-+%5Cphi%7D%7B%5Ckappa%7D%5Cright%5D+%5Cleft%28N+%2B+1+-+N%5E%2A%5Cright%29%7D&bg=f5efe0&fg=000000&s=0&c=20201002)

For large 

which is the number of surviving species.
Clearly, this is non-zero for a finite catalytic rate 
These computations clearly should be taken with a grain of salt. As pointed out in several sources, hypercycles describe closed systems, but, life exists in an open system driven by an energy flux. But, the interesting thing is, despite this, the very last calculation shows that there is clear division between molecules 

2 replies on “Mathematical Origins of Life”
Reblogged this on scyteck.
Greaat reading your post